During my undergrad, I came across the topic Circadian Clocks a lot and I had a special interest in ryegrass. Ryegrass is a forage and turf grass species found in temperate regions it is commonly the most dominant grass species. Perennial ryegrass is grown in the USA, China, Japan, UK, Australia and accounts for 95% of forage grass in Ireland (Teagasc). This led me to look into plants and Circadian Clocks more. Below is a bit I did on Circadian Clocks whilst reading on the topic.
Plants have adapted to seasons using a mechanism that involves photoperiodic flowering. Plants respond to photoperiods in two main categories, plants that flower under longer photoperiods which are long-day plants. These plants require less darkness to induce flowering (eight hours of darkness). Short day plants require less than 12 hours of daylight. These plants bloom in the spring or later in the fall. Some plants are not responsive to changes in photoperiod, and these are day-neutral plants like sunflowers (Helianthus annuus) and peas (Pisum sativum). Molecular analysis of Arabidopsis thaliana has exposed on a molecular level how day length in photoperiodic flowering is measured by long-day plants (Izawa 2007). Ryegrass needs vernalisation (Vernalization is the induction of a plant’s flowering process by exposure to the prolonged cold of winter) in order to flower (Pyo et al 2014)
These biological events are timed correctly due to the circadian clocks synchronising with the day-night cycle and seasons. (Terzibasi-Tozzini et al 2017). This circadian regulation is important in plants since it adjusts several processes such as hormonal signals, growth, development, internal metabolic and stomatal opening to 24h cycles. The circadian clock is a biological oscillator that involves the transcriptional and translational feedback loops resulting in a “c. 24 h” rhythm (Harmer, 2009). The clock interacts with the constantly changing environment to which plants need to adapt to daily. Photoperiod and temperature are the biggest environmental factors that entrain the circadian regulation (Hirschie et al., 2003). It is widely thought that the circadian clock has evolved towards a more complex and thereby flexible and robust process which helps plants to cope with irregular variations in sunlight and temperature (Rand et al 2004). The circadian clock synchronises and controls the necessary cellular response to anticipate the shift from light and dark (5). This timekeeper allows plants to adapt to seasons mostly through gene expression and protein activity (Dodd et al 2015).
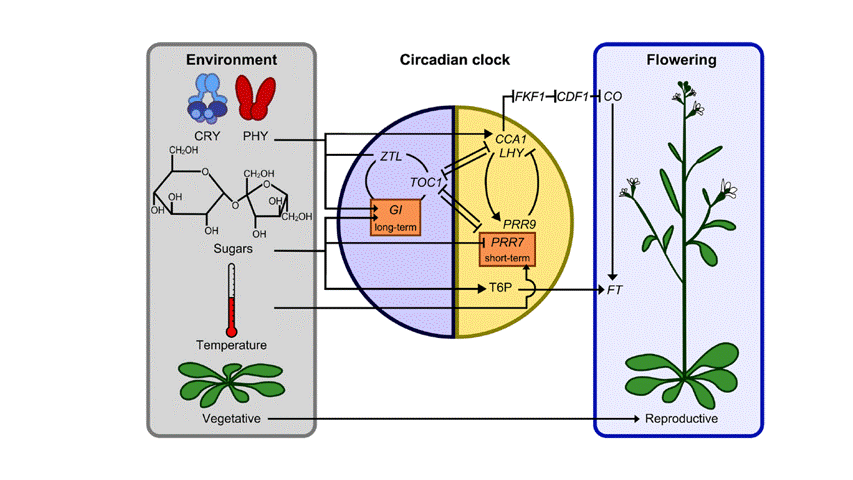
Fig.1 Interactions between sugar and light signals, the circadian oscillator, and the photoperiodic regulation of flowering (Dodd et al 2015).
Haydon et al experimentally demonstrated that the circadian clock is also entrained by sucrose. They also proposed that low sugar increased the expression of certain clock components such as PRR7 (proline rich 7), suggesting that PRR7 is necessary for entrainment to sugars (Haydon et al 2013; Dodd et al 2015). Figure.1 shows the relationship between the signals of sugar and light, the circadian oscillator, and the photoperiodic regulation of flowering. Photoreceptors detect the low-intensity light at dawn, which will then entrain the circadian oscillator. When light intensity increases later in the morning, a second entrainment event occurs. The oscillator is advanced in the response to the rhythmic accumulation of sugars that occurs due to the daily stimulation of photosynthesis every morning (Seo and Mas 2015).
Here are two interesting papers I read recently
- Sinturel, F., Petrenko, V. and Dibner, C., 2020. Circadian clocks make metabolism run. Journal of molecular biology.
- Segers, A. and Depoortere, I., 2021. Circadian clocks in the digestive system. Nature Reviews Gastroenterology & Hepatology, pp.1-13.
References
Dodd, A.N., Belbin, F.E., Frank, A. and Webb, A.A., 2015. Interactions between circadian clocks and photosynthesis for the temporal and spatial coordination of metabolism. Frontiers in plant science, 6, p.245.
Gilliland, T.J., Barrett, P.D., Mann, R.L., Agnew, R.E. and Fearon, A.M., 2002. Canopy morphology and nutritional quality traits as potential grazing value indicators for Lolium perenne varieties. The Journal of Agricultural Science, 139(3), pp.257-273.
Harmer, S.L. and Kay, S.A., 2005. Positive and negative factors confer phase-specific circadian regulation of transcription in Arabidopsis. The Plant Cell, 17(7), pp.1926-1940.
Harmer, S.L., 2009. The circadian system in higher plants. Annual review of plant biology, 60.
Haydon, M.J., Mielczarek, O., Robertson, F.C., Hubbard, K.E. and Webb, A.A., 2013. Photosynthetic entrainment of the Arabidopsis thaliana circadian clock. Nature, 502(7473), p.689.
Hirschie Johnson, C., Elliott, J.A. and Foster, R., 2003. Entrainment of circadian programs. Chronobiology international, 20(5), pp.741-774.
Izawa, T., 2007. Daylength measurements by rice plants in photoperiodic short‐day flowering. International review of cytology, 256, pp.191-222.
Pyo, Y., Park, S., Xi, Y. and Sung, S., 2014. Regulation of flowering by vernalisation in Arabidopsis. In Advances in Botanical Research (Vol. 72, pp. 29-61). Academic Press.
Rand, D.A., Shulgin, B.V., Salazar, D. and Millar, A.J., 2004. Design principles underlying circadian clocks. Journal of The Royal Society Interface, 1(1), pp.119-130.
Seo, P.J. and Mas, P., 2015. STRESSing the role of the plant circadian clock. Trends in plant science, 20(4), pp.230-237.
Terzibasi-Tozzini, E., Martinez-Nicolas, A. and Lucas-Sánchez, A., 2017, October. The clock is ticking. Ageing of the circadian system: from physiology to cell cycle. In Seminars in cell & developmental biology (Vol. 70, pp. 164-176). Academic Press
Vogel, J. and Hill, T., 2008. High-efficiency Agrobacterium-mediated transformation of Brachypodium distachyon inbred line Bd21-3. Plant cell reports, 27(3), pp.471-478.